






















































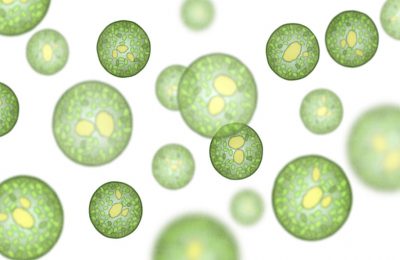
























































IntroduzioneLa vita sembra essersi originata nelle acque dei primordiali oceani. Dopo aver sperimentato diverse strategie di nutrizione e diverse organizzazioni cellulari, gli esseri viventi si sono perfezionati ed hanno iniziato quel lungo cammino che li ha portati fino a noi. La cellula eucariote ed un metabolismo basato su processi ossidoriduttivi, quali la respirazione cellulare e la fotosintesi, hanno permesso ai primi semplici organismi di evolvere fino ad originare forme sempre più complesse e meglio adattate alle esigenze imposte dalla colonizzazione dell’ambiente Terra, a cominciare dai mari.Caratteristiche generali Negli ambienti acquatici, sia d’acqua dolce che salata e anche negli ambienti molto umidi sulla terraferma, vive il gruppo delle Alghe nel quale è possibile individuare le tappe seguite dall’evoluzione. Tale gruppo, infatti, non è da considerarsi una vera e propria categoria tassonomica, in quanto in essa confluiscono organismi che presentano numerose dif ferenze e vengono classificati in divisioni fra loro anche molto distanti. E’ comunque rimasta la consuetudine di indicare col nome di Alghe tutti quei vegetali acquatici, sia unicellulari che pluricellulari, che hanno una organizzazione del corpo molto semplice, anche se a volte simile a quella delle piante superiori come aspetto esterno. Nella loro struttura non si trovano mai tessuti specializzati ad una determinata funzione, nè organi come radici, fusto, foglie, ma ciascuna delle loro cellule è in grado di svolgere la totalità delle reazioni metaboliche necessarie alla vita. Le alghe unicellulari Si può pensare che le Alghe unicellulari siano i rappresentanti più antichi di questo gruppo, o almeno quelli che hanno mantenuto più di altri le caratteristiche presenti nei primi tipi di Alghe.
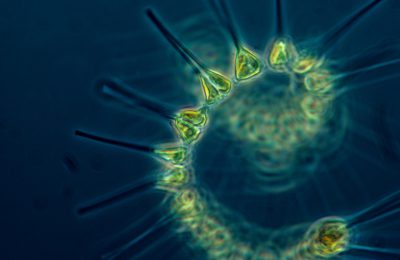
Euglenofite Un indice di primitività è fornito dalla divisione delle Euglenofite. Queste Alghe unicellulari sono prive di parete cellulare cellulosica, ma il loro corpo è circondato da uno strato elastico e molle detto periplasto, originato dall’indurimento della parte più periferica del citoplasma. Possono presentare dei cloroplasti, i quali contengono sempre clorofilla a, e raramente b, oltre al P-carotene e xantofille. Molte sono eterotrofe, parassite o saprofite. Alcune specie di Euglene autotrofe, in condizioni di buio persistente, perdono i loro cloroplasti e passano a nutrirsi per via eterotrofa. Questa caratteristica induce a pensare che esse possano essere molto antiche, risalenti ai primi tempi dell’evoluzione, quando i viventi erano prevalentemente eterotrofi, ma capaci anche di fotosintesi. Altra caratteristica a sostegno di questa ipotesi è il fatto che esse presentano una divisione cellulare molto primitiva, mancante di fuso e anomala sotto vari aspetti. La sostanza alimentare di riserva non è l’amido, ma il paramylon, un altro polisaccaride insolubile in acqua, tipico di molte Alghe, anche superiori. Le Euglene si muovono grazie ad un lungo flagello posto anteriormente e che trascina l’organismo (altro fatto strano dato che i flagelli in genere spingono l’organismo che li porta). La riproduzione avviene tipicamente per scissione longitudinale, mentre molto rara è la riproduzione sessuale. Vivono in ambienti di acqua dolce e spesso colorano le acque in modo particolare, data la ricchezza di pigmenti che a volte si accumulano entro il citoplasma. Pyrrofite Altra divisione che presenta caratteri di primitività, è quella delle Pyrrofite che, come le Euglenofite, sono, per certi aspetti, una via di mezzo fra le Piante e gli Animali. Le Pirrofite sono nella maggior parte flagellati unicellulari. Alcune sono autotrofe, altre eterotrofe, per cui i cloroplasti e la parete cellulare non sono sempre presenti. Nei cloroplasti sono contenuti sempre clorofilla a e c, betacarotene e vari tipi di xantofille mentre la parete cellulare è formata da cellulosa. Essa è tipicamente divisa in due o più parti (valve), che delineano i due solchi, uno trasversale e l’altro longitudinale, entro i quali sono compresi i due flagelli interessati al movimento. Nel nucleo sono presenti cromosomi primitivi, tipicamente privi di istoni. Le riserve alimentari risultano costituite da amidi. Anche in queste Alghe la riproduzione avviene prevalentemente per via asessuale, raramente sessuale. Sono abbondanti nelle acque, dove periodicamente vanno incontro a cicli di crescita incontrollata per cui aumentano di numero vertiginosamente, provocando morie estese di pesci, a causa di sostanze tossiche che accumulano nel loro corpo oppure per sottrazione di ossigeno dalle acque. Le ‘fioriture’ delle Pirrofite sono facilmente individuabili dato il colore rosso cupo assunto dalle acque a causa dei pigmenti presenti nel corpo di tali Alghe Crisofite Sono Alghe unicellulari o coloniali filamentose molto semplici. In esse la parete presenta a volte cellulosa, mentre altre volte è fortemente silicizzata e divisa in due valve finemente scolpite, come nelle diatomee. A volte sono presenti i cloroplasti che contengono tipicamente clorofilla a e, raramente, c oltre ai soliti pigmenti accessori. In queste Alghe la divisione mitotica comincia ad essere simile a quella degli eucarioti superiori ed anche la riproduzione sessuale inizia ad apparire con una certa frequenza e con meccanismi che denotano un evidente cammino evolutivo. Clorofite La divisione delle Clorofite presenta gruppi sistematici nei quali si può notare un passaggio graduale dalla unicellularità alla pluricellularità. Sono tutte dotate di clorofilla a e b e di carotenoidi in quantità simili alle Piante terrestri; proprio per questo sono considerate loro progenitrici. Nelle forme più semplici si nota una organizzazione cellulare simile a quella delle Euglene, con flagelli che permettono la mobilità e riproduzione generalmente asessuata. Tipico rappresentante di questo gruppo è Chlamidomonas, nel quale però si assiste sempre ad una riproduzione sessuata con produzione di uno zigote che, dopo un periodo variabile di mobilità, si circonda di una spessa parete per potere resistere durante la cattiva stagione. Alcune specie di organismi con organizzazione simile a Chlamidomonas vivono in colonie sferiche od ovoidali cave, composte da numerosi individui (fino a 50.000 o più) separati fra loro o collegati da sottili filamenti citoplasmatici. Questa condizione ricorda quindi molto da vicino la pluricellularità. In ogni caso nelle colonie giovani tutte le cellule sono perfettamente uguali fra loro. Tipico esempio è dato dal genere Volvox, organismo flagellato coloniale che vive nelle acque dolci. Quando la colonia invecchia alcune cellule si specializzano per la riproduzione e si trasformano in gonidi, grosse cellule interne, le quali potranno sviluppare nuove colonie sia sessualmente che asessualmente. La riproduzione sessuata, si verifica in prossimità della cattiva stagione e dà origine ad uno zigote quiescente.
Alghe pluricellulari Struttura La pluricellularità delle Alghe non deve essere confusa con quella delle piante superiori, infatti in esse non compare quasi mai specializzazione funzionale delle cellule, e quindi non si trovano mai tessuti veri e propri. Molto spesso l’aspetto pluricellulare viene acquisito solo perché le singole cellule dopo essersi originate in seguito a mitosi non si separano, ma restano unite per mezzo di sostanze pectiche che impregnano le pareti, oppure perché la separazione fra le cellule figlie è incompleta. Nel primo caso si ottengono in genere degli individui che in realtà hanno più carattere coloniale che pluricellulare, ed il loro aspetto è in genere filamentoso perché i fusi mitotici sono tutti orientati nello stesso verso. Molto spesso i filamenti così ottenuti si intrecciano fra loro dando origine ad una sorta di feltro che è, in realtà, un falso tessuto, anche se a volte il suo aspetto può indurre in errore. In questi casi, infatti, l’Alga assume una struttura che può ricordare un cormo, date le ampie lamine simili a foglie (filloidi) ed i prolungamenti basali simili a radici (rizoidi), oltre a strutture fluttuanti ramificate simili a fusti (cauloidi).
Riproduzione Il lungo cammino evolutivo percorso dalle Alghe è testimoniato dalle modalità con le quali esse si riproducono. Nelle Alghe pluricellulari, la riproduzione può essere sia asessuata che sessuata, ma quest’ultima ha generalmente maggior importanza pratica. La riproduzione asessuata può verificarsi, a volte, per semplice frammentazione del tallo, oppure per produzione di cellule specializzate che si staccano dalla pianta madre ed acquistano una vita indipendente (spore). La riproduzione sessuale, che si attua per mezzo della fusione di due gameti, è molto sviluppata e può verificarsi in modi e forme diverse, rappresentative del grado di evoluzione raggiunto dalla specie. Si può infatti passare da fenomeni di isogamia, nei quali i due gameti sono mobili e di identica forma, a fenomeni di anisogamia, in cui incomincia a notarsi il differenziamento fra gamete femminile, più grosso, e gamete maschile, più piccolo, nonostante entrambi mantengano la propria mobilità. Il passo successivo si nota con la oogamia, nella quale il grosso gamete femminile è divenuto immobile, trasformandosi in oosfera, mentre il gamete maschile mantiene la caratteristica mobilità. I gameti di sesso diverso si formano generalmente su tali separati, raramente sullo stesso. Nelle Alghe pluricellulari si incontra, inoltre, per la prima volta, una regolare alternanza fra riproduzione asessuata e riproduzione sessuata, che si susseguono nel corso del tempo originando particolari cicli metagenetici. A seconda che in questi cicli prevalga la generazione diploide o quella aploide si parlerà di ciclo diplonte o aplonte rispettivamente, oppure di ciclo aplo-diplonte qualora le due generazioni siano equivalenti. Anche nei cicli metagenetici delle Alghe si può trovare testimonianza di una evoluzione riproduttiva che avrà il suo seguito nelle Piante superiori. Ciclo Aplonte Il ciclo più primitivo è considerato quello aplonte che si ritrova tipicamente nel genere Oedogonium, un’Alga filamentosa con tallo non ramificato, che vive in stagni e corsi d’acqua. Molte specie sono eterotalliche, cioè il tallo maschile e quello femminile sono distinti, a volte anche diversi come aspetto. Ciclo aplo-diplonte Nel genere Ulva si ha invece un tipico ciclo aplo-diplonte, in cui lo sporofito ed il gametofito si equivalgono. Ulva è un’Alga comune, che cresce in acqua marina lungo il margine fra alta e bassa marea. Ciclo diplonte In altri generi, come ad esempio in Fucus, il gametofito non esiste più e l’unico momento aploide del ciclo è dato dalle spore, che funzionano quindi anche da gameti. Si ha quindi un ciclo chiaramente diplonte. Metabolismo Le Alghe pluricellulari sono tutte obbligatoriamente autotrofe per fotosintesi clorofilliana, ed in questo si dimostrano quindi organismi già piuttosto evoluti rispetto ad alcune Alghe unicellulari, per le quali è ancora possibile l’alternativa eterotrofa. Esse contengono tutte clorofilla a e b ed una diversa quantità di pigmenti accessori tipici di ogni gruppo, che viene utilizzata quale criterio di classificazione. Nutrizione minerale Le Alghe, rispetto alle piante terrestri, risultano notevolmente avvantaggiate dal fatto di vivere immerse in acqua. Infatti l’acqua fornisce sostegno, per un noto principio di fisica, e protezione dalla siccità. Proprio per questi motivi esse non hanno sviluppato tessuti necessari a sostenerle, nè cuticole protettive. Tutte le cellule sono immerse nel medesimo mezzo e quindi non sono necessari neppure i tessuti di trasporto dei sali minerali e dell’anidride carbonica, che si ritrova disciolta in grandi quantità nelle acque salate. N.B. Dal momento che il modo con cui avviene la nutrizione minerale ed idrica delle Alghe è lo stesso che si presenta anche nelle Piante terrestri, si tratteranno in questa sede in modo dettagliato i fenomeni fisici che regolano l’assorbimento, richiamando le conoscenze di biologia generale già in vostro possesso. Trasporto La membrana cellulare, come visto nel precedente capitolo, è dotata della proprietà di semipermeabilità, cioè di selettività: solo alcune sostanze sono libere di muoversi attraverso il suo spessore, per le altre il passaggio è condizionato al verificarsi di particolari fenomeniUno di tali fenomeni è il trasporto passivo. Esso si verifica quando a livello della membrana sono presenti delle particolari molecole proteiche in grado di riconoscere la sostanza e di farla passare attraverso lo spessore della membrana, in seguito ad un cambiamento nella loro configurazione spaziale. Quando la concentrazione ai due lati della membrana è diventata le stessa, il fenomeno si arresta e non si ha più passaggio netto della sostanza.In particolari casi però la cellula potrebbe aver bisogno di una sostanza in quantità maggiori rispetto a quelle raggiungibili con il semplice- bilanciamento delle concentrazioni. In questo caso essa deve accumulare al suo interno, o eliminare completamente all’esterno, la sostanza in oggetto. Ciò è possibile ricorrendo al fenomeno del trasporto attivo, durante il quale la cellula spende enormi quantità di energia: la sostanza, infatti, viene trasferita dal compartimento in cui è presente in minor concentrazione, verso quello dove la sua concentrazione è maggiore, in contrasto con le leggi della diffusione. Anche in questo caso, a livello di membrana, deve essere presente una molecola trasportatrice, ma stavolta essa deve svolgere funzioni anche enzimatiche, deve cioè scindere grandi quantità di ATP, per poter ottenere l’energia sufficiente a permettere il fenomeno. Osmosi A volte la membrana si mostra completamente impermeabile a determinate sostanze e per loro non esiste possibilità alcuna di attraversarla. In questo caso non sarà la sostanza a muoversi, ma l’acqua in cui essa è disciolta. Tutte le sostanze nutritive sono, infatti, sempre presenti in soluzione: esse formano il soluto, mentre l’acqua è il solvente. Ora, per equilibrare la concentrazione di due soluzioni, si può agire in due modi; o si fa passare da una soluzione all’altra l’esatta quantità di soluto che compensi il divario fra le due concentrazioni, oppure si trasferisce acqua, cioè solvente, dal compartimento in cui la concentrazione della sostanza è minore (notare che di acqua ce ne sarà di più) verso quello dove la concentrazione è maggiore (qui di acqua ce n’è di meno perché c’è più soluto). Al passaggio di acqua attraverso una membrana semipermeabile viene dato il nome di osmosi. Nei vegetali acquatici i fenomeni osmotici sono la norma, in quanto essi sono completamente circondati da soluzioni di composizione sempre variabile. Se la soluzione esterna alla cellula ha una concentrazione di soluti maggiore del citoplasma cellulare (se cioè è ipertonica), l’acqua tenderà ad uscire dalla cellula per cercare di diluire la soluzione; se invece la soluzione ha una concentrazione minore rispetto alla cellula, l’acqua entra nella cellula stessa. In quest’ultimo caso se la differenza di concentrazione è alta, enormi quantità d’acqua entrerebbero nella cellula, provocandone una dilatazione ed infine la distruzione. Nelle cellule vegetali tale evento non si verifica grazie alla parete esterna di cellulosa, che a causa della sua rigidità impedisce l’eccessivo dilatarsi della cellula. L’acqua che entran seguito ad un fenomeno di osmosi si accumula entro il vacuolo centrale per cui, ai fini di questo fenomeno, la membra- na veramente importante à il tonoplasto. L’acqua accumulata nel vacuolo, genera una pressione idrostatica rivolta verso la parete, che limita, fino ad impedirlo completamente, il movimento dell’acqua. A tale pressione viene dato il nome di pressione osmotica ed il suo valore può variare moltissimo, arrivando fino a 20 atmosfere. Gli organismi che vivono in acque dolci, a basso contenuto salino e quindi ipotoniche, sono soggetti ad un notevole flusso di acqua in entrata verso il vacuolo. Per limitare il fenomeno ed impedire l’esplosione della cellula, gli organismi debbono potenziare la propria parete, rendendola più rigida per mezzo della impregnazione con sali inorganici, oppure devono evolvere meccanismi di eliminazione dell’acqua in eccesso. A questo scopo la maggior parte delle Alghe unicellulari è dotata di strutture particolari dette vacuoli contrattili. Al contrario, le Alghe che vivono in acque ipertoniche, con molti sali, sono soggette ad una continua perdita di acqua. Per evitare ciò, esse accumulano soluti entro il vacuolo in modo da innalzare la propria concentrazione salina. Pressione di turgore L’acqua presente nel vacuolo di ogni cellula vegetale genera una pressione osmotica rivolta contro le pareti della cellula, definita pressione di turgore e determina in tal modo il turgore cellulare. PH20 = PT–P0 dove PH20 corrisponde al potenziale d’acqua, PT alla pressione di turgore e PO alla pressione osmotica. PO è un valore teorico, dipendente dalle concentrazioni dei soluti e quindi non sarà mai possibile che in condizioni reali esso possa essere eguagliato; ciò determina un potenziale d’acqua effettivo anche in condizioni di apparente equilibrio osmotico. Classificazione Le Alghe pluricellulari vengono classificate in base al tipo di pigmenti accessori e quindi in base al loro colore. Si distinguono in tre grosse divisioni: Clorofite, Feofite e Rodofite. Clorofite Le Clorofite sono dette anche Alghe Verdi per il colore verde vivo dei cloroplasti, dovuto alla presenza di clorofilla a e b e di carotenoidi, nelle stesse proporzioni delle Piante terrestri. Questo fatto ha indotto a pensare che la conquista del mondo subaereo sia iniziata proprio a partire da qualche gruppo ancestrale di queste Alghe. In questa divisione oltre alle forme pluricellulari, entro le quali non si nota mai una grande differenziazione funzionale delle cellule, sono presenti anche diverse forme unicellulari.
Neofite La divisione delle Feofite comprende le forme di maggiori dimensioni che possono raggiungere anche i 100 metri di lunghezza. Sono caratterizzate da un’ intensa colorazione bruna dovuta alla presenza, oltre che di clorofilla a e c , di beta-carotene, di fucoxantina ed altre xantofille in grandi quantità, tali da giustificare il nome di Alghe Brune. Vivono generalmente in ambiente marino, in zone temperate o fredde, dove crescono in tale quantità da formare vere e proprie praterie sommerse, come nel Mar dei Sargassi. Rodofite La divisione delle Rodofite comprende forme generalmente pluricellulari, di dimensioni solitamente ridotte, con differenziazione funzionale molto scarsa. Sono caratterizzate da presenza di clorofilla a e d, oc e beta-carotene, luteina e ficoeritrina; sono dette Alghe Rosse per il loro tipico colore e vivono in ambienti marini spingendosi fino a 200 m di profondità. Non possiedono forme flagellate e accumulano le sostanze di riserva in uno speciale polisaccaride detto amido delle floridee
Ambiente marino I diversi pigmenti presenti hanno una importante funzione fisiologica, in quanto permettono alle alghe di catturare efficacemente la luce solare che filtra attraverso l’acqua. Le Clorofite sono destinate ad occupare lo strato più superficiale dei fondali, quello vicino alla linea di demarcazione fra ambiente aereo e acquatico, in quanto la composizione dei loro pigmenti permette di assorbire efficacemente solo le radiazioni che per prime vengono eliminate dallo spettro solare, e cioè quelle rosse. Le alghe brune, grazie al loro colore possono invece spingersi più in basso, fino a 100 metri se l’ acqua è limpida. Spesso esse si ancorano al fondo con un potente rizoide e poi si spingono con il resto del tallo fino alla superficie, formando dei fitti ammassi. Le Rodofite, grazie alla particolare combinazione di pigmenti, possono spingersi invece a profondità maggiori, anche più di 200 metri in caso di acque limpide, ove giungono solo le radiazioni verdi e azzurra. |
La vita nelle acque
3297








TAGalghe



